Paleocene mammals of the world
by Martin Jehle
http://www.paleocene-mammals.de/
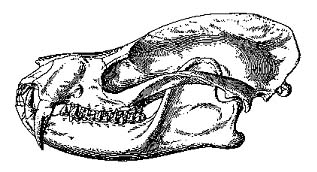 |
Figure 1. Skull of Deltatherium fundaminis,
one of the first Paleocene mammals described from North America. Its relationships are
still debated. From Williamson (1996). For a reconstruction, see the figure below -- it is
climbing down a branch near the top center. |
The first 10 million years of the age of mammals
Mammals appear first in the late Triassic, at about the same time as dinosaurs.
Throughout the Mesozoic, most mammals were small, fed on insects and lead a nocturnal
life, whereas dinosaurs were the dominant forms of life on land. This situation changed
abruptly about 65 million years ago, when dinosaurs disappeared with the exception of
their descendants, the birds, probably as the result of a meteorite impact. The epoch
right after the mass extinction of the dinosaurs is called the Paleocene. At the beginning
of the Paleocene epoch, the world was practically without larger sized terrestrial
animals. This unique situation was the starting point for the great evolutionary success
of the mammals. Only ten million years later, at the end of the Paleocene, they had
occupied a large part of the vacant ecological niches. By this time, the landscape was
teeming with small insectivorous and rodent-like mammals, medium sized mammals were
searching the forests for any kind of food they could cope with, the first large (but not
yet gigantic) mammals were browsing on the abundant vegetation, and carnivorous mammals
were stalking their prey.
Despite this impressive diversification, most Paleocene mammals are still on a
primitive level of anatomy in comparison to mammals of today. Often they show only the
first beginning of specializations that characterize their descendants from later epochs,
such as optimization of the teeth for a special kind of food or adaptations of the limbs
to fast running. The Paleocene mammalian fauna is therefore often called archaic. The
beginning of the following epoch, the Eocene, brought about an important modernization of
this fauna. Several groups of mammals with more modern appearance spread over the northern
hemisphere at this time, whereas the decline of the archaic forms started.
The Paleocene is a crucial time in the history mammals. Unfortunately, mammal fossils
from this epoch are either scarce or entirely unknown in many parts of the world. Thus we
can only speculate how the fauna of whole continents looked after the extinction of
dinosaurs. Even where fossils occur, most species are only known from their characteristic
teeth, and skeletons are only known for a few forms. Nevertheless, knowledge of Paleocene
mammals is steadily increasing. This website tries to summarize what we know about these
first ten million years of the age of mammals.
Introduction to Paleocene mammals
The target of this section of the site is to compile the most interesting facts about
Paleocene mammals. Paleontological literature can be a somewhat dry matter, especially if
the subject is known from not more than fossil teeth and the entire animal remains largely
unknown, as is often true for early mammals. Naturally, biological interpretations are
quite limited in such cases. Nevertheless, there are exceptional cases where we can get a
more complete idea of the animal and its behavior. I have tried to extract from the
literature these highlights, besides giving an overview of the evolution and relationships
of Paleocene mammals.
 |
| Reconstruction of a middle Paleocene habitat in New Mexico, USA.
On the forest floor two mesonychids of the genus Ankalagon are eating the small
crocodile Allognathosuchus. On the left the coati-like arctocyonid Chriacus is
skillfully climbing up a plane-tree and is watching a Deltatherium on the small
branch in the middle. Ankalagon and Chriacus are primitive members of the
hoofed mammals, whereas the relationships of Deltatherium are still debated.
Drawing by Gregory S. Paul, from Stanley (1989). |
Multituberculates:
Heyday of the longest lived mammalian order
The order Multituberculata, informally also known as 'multis', is a diverse lineage of
Mesozoic to early Cenozoic mammals that occupied a rodent-like niche. They appear first in
the late Jurassic and are last known from the early Oligocene. The late Triassic
haramiyids were sometimes considered as early multituberculates. More complete fossils
have recently shown that haramiyids are a very different group of early mammals. But even
without haramiyids, multituberculates existed for a time span of about 100 million years,
the undisputed record for an order of mammals.
Multituberculates do not belong to any of the groups of mammals living today: the
primitive egg-laying monotremes and the more advanced marsupials and placentals, both also
known as therians. The relationships of the multituberculates to these groups are still
debated. Multituberculates have been considered as either a group that branched off even
before the monotremes, as close relatives of the monotremes or as sister group of the
therian mammals. Anyway, multituberculates were clearly very mammal-like, both in details
of the internal anatomy, like the structure of the middle ear with the three auditory
ossicles, and in external appearance, like the recently demonstrated possession of hair
(see below). The anatomy of the pelvis suggests that multituberculates did not lay eggs
like monotremes but gave birth to very small, immature young like marsupials.
In the late Cretaceous multituberculates were widespread and diverse in the northern
hemisphere, making up more than half of the mammal species of typical faunas. Although
some lineages became extinct during the faunal turnover at the end of the Cretaceous,
multituberculates managed very successfully to cross the K/T boundary and reached their
peak of diversity during the Paleocene. They were an important component of nearly all
Paleocene faunas of Europe and North America, and of some late Paleocene faunas of Asia.
Multituberculates also were most diverse in size during the Paleocene, ranging from the
size of a very small mouse to that of a beaver.
 |
| Figure 1: Reconstruction of the Paleocene multituberculate Ptilodus,
about 50 cm in length. From Cox (1998). |
Ptilodus is a typical medium-sized multituberculate from the Paleocene of North
America. It is known from a nearly complete skeleton discovered in Canada, which shows
that Ptilodus can best be compared with today's squirrels. The skeleton of Ptilodus
shows several adaptations for life in trees, for instance sharp claws. Like in squirrels,
the foot of Ptilodus was very mobile and could be reversed backward, which allowed
the animal to climb down trees with its head pointing downward. A marked difference from
squirrels is the long prehensile tail, which Ptilodus used like a fifth limb when
climbing in trees.
The dentition of Ptilodus and other multituberculates is most similar to rodents
in the enlarged front teeth, especially the pair of lower incisors, which are followed by
a gap (called diastema) in the lower jaw. The last lower premolars of most
multituberculates form large, serrated blades. This type of teeth is called plagiaulacoid
after the Mesozoic multituberculate genus Plagiaulax. Several groups of mammals
have independently developed plaugiaulacoid teeth, although not to the same degree as
multituberculates like Ptilodus:
- an early Tertiary group of primates called carpolestids
- several extinct South American marsupials like the early Tertiary polydolopids
- the recent rat-kangoroos of Australia
- the recent pygmy possum Burramys (another Australian marsupial)
The back of the multituberculate jaw was occupied by a battery of molars that formed a
grinding mechanism. These molars carried several longitudinal rows of many small cusps (or
tubercles), which is the origin of the name Multituberculata.
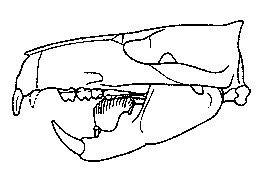 |
Figure 2: Skull of Ptilodus montanus from the middle
Paleocene of Montana, U.S.A, showing the blade-like lower premolar. After Miao (1993). |
The diet of multituberculates is a long debated issue. Following the analogy to
rodents, it could be assumed that multituberculates were herbivores. However, even today's
rodents do not all feed on vegetable material only. In addition, the blade-like lower
premolar must be taken into account. The recent rat-kangaroos, which share this feature,
include not only herbivores but also omnivores that feed on plants, insects or even
carrion. A similar diet can be imagined for multituberculates, too. In this scenario, the
enlarged incisors would be have been used for picking up and killing insects or other
prey. The blade-like premolars could have served both for biting hard shelled seeds and
for chopping up small prey.
Members of a specialized family of multituberculates, the Taeniolabididae, can be
considered more confidently as herbivores. Like rodents, taeniolabidids (and some related
multituberculate families) had developed gnawing teeth with a self-sharpening mechanism.
Only the front side of the incisors is covered with hard enamel. The rest of the tooth
consists of softer material which is worn away more easily. Therefore the tooth has always
a sharp cutting edge at the front. The blade-like premolars are strongly reduced, whereas
the molars form enlarged, complex grinding surfaces with an increased number of cusps. The
type genus of the family is Taeniolabis from the early Paleocene of North America.
This beaver-sized animal is the largest known multituberculate. Taeniolabis had a
massive skull, low in profile, with a short, blunt snout. Its well-developed gnawing teeth
and complex molars are strong evidence that Taeniolabis, unlike most other early
Paleocene mammals, had specialized on vegetable diet.
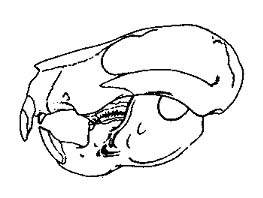 |
Figure 3: Skull of Taeniolabis taoensis from the early
Paleocene of New Mexico, U.S.A, about 180 mm in length. Note development of gnawing teeth.
From Sloan (1981). |
Remarkable fossils from the late Paleocene of China are known for a second
taeniolabidid, Lambdopsalis. The skeleton of Lambdopsalis shows
specializations for digging, so this multituberculate was probably living in burrows.
Well-preserved skulls indicate that the ear of Lambdopsalis was inefficient for
hearing high-frequency airborne sound, but well-adapted for hearing low-frequency
vibrations, which would be useful for a burrowing animal. Lambdopsalis was
obviously a favorite prey of carnivorous mammals, as evidenced by its occurrence in
coprolites (fossilized excrements) of carnivores at a Chinese locality. Absolutely unique
is the fact that these coprolites preserve fossilized hair of the prey, including that of Lambdopsalis,
which is very similar in structure to hair of living mammals. This is the only direct
evidence that the extinct multituberculates had hair, and at the same time the oldest
known occurrence of fossil mammalian hair.
After the diversity of multituberculates had reached a maximum in the Paleocene, it
started to decline towards the end of this epoch. This may be due to competition with an
increasing number of placental herbivores, especially with the archaic hoofed mammals
called condylarths and with primates. In the latest Paleocene true rodents appeared as
additional competitors that were particularly close in ecology. A number of
multituberculate genera survived into the Eocene, but few new genera developed, and
multituberculates became extinct in the early Oligocene.
An odd group of late Mesozoic to early Cenozoic mammals from the ancient landmass of
Gondwana is worth mentioning here since it may represent a southern radiation of
multituberculates. These poorly known animals, appropriately called gondwanatheres, were
first found in the late Cretaceous (Gondwanatherium, Ferugliotherium) and early
Paleocene (Sudamerica) of South America. Rare fossils have recently also turned up
in the Late Cretaceous of Madagascar and India, all former parts of disintegrating
Gondwana. At least Gondwanatherium and Ferugliotherium had enlarged incisors
for gnawing. But the most striking feature of advanced gondwanatheres like Gondwanatherium
and Sudamerica are the very high-crowned molars. In the later Tertiary many
groups of plant-eating mammals have developed high-crowned teeth to cope with the
spreading grasses that strongly wear down teeth. However, such teeth are unique for the
Mesozoic and the earliest Cenozoic, and we do not know what triggered their early
evolution in gondwanatheres. Grasses did not appear in South America until much later, and
fossils of Sudamerica were probably deposited in a mangrove swamp.
Some gondwanatheres were previously considered as early member of the Xenarthra
(armadillos, anteaters and sloths) due to somewhat similar teeth and their geographic
occurrence in South America, the home of the xenarthrans. Detailed studies of the
dentition, including the microstructure of the enamel covering the teeth, have suggested
that gondwanatheres may instead be the first 'multis' known from southern continents. More
complete fossils will hopefully resolve the debate on the relationships of this enigmatic
group from Gondwana.
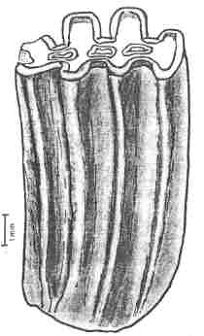 |
Figure 4: High-crowned tooth of the gondwanathere Sudamerica
ameghinoi from the early Paleocene of Patagonia, Argentina. vertical bar to the left
is 1mm in length; the tooth is about 8mm high.The From Scillato-Yané & Pascual
(1985). |
Marsupials: A southern success story
Marsupials are often associated with the Australian continent where such popular
animals as kangaroos and koalas live. Less widely known is that a number of marsupials
occur in South America today. These include the opossums, the only marsupials that also
occur in the northern hemisphere today.
Surprisingly, Mesozoic marsupials are mainly known from North America, and they may
have originated there during the Cretaceous. Primitive opossum-like marsupials are among
the most common and most diverse mammals in the late Cretaceous of North America. They
include cat-sized forms like Didelphodon, one of the largest Mesozoic mammals - and
one of the most widely known thanks to its appearance in "Walking with
Dinosaurs". Didelphodon had teeth specialized for crushing, and it has been
suggested that it fed on hard shelled food like clams, snails, turtles or even young
ankylosaurs.
The diversity of North American marsupials was dramatically reduced at the end of the
Cretaceous. Only one single lineage, represented by late Cretaceous Alphadon and
Paleocene Peradectes, survived the faunal turnover. Thus marsupials fared hardly
better than dinosaurs in North America, and worse than reptiles on the whole. Peradectes
survived into the Eocene, and a few other opossum-like marsupials appear in North America
during the Paleocene. However, marsupials never became a significant component of northern
faunas again and finally became extinct there. Opossums reentered North America from South
America only a short time ago, after the Isthmus of Panama had formed in the Pliocene.
 |
| Figure 5: Reconstruction of Late Cretaceous Alphadon, based
mainly on the appearance of today's opossums since only the dentition of this animal is
known. This reconstruction should be equally valid for Peradectes, the Paleocene to
Eocene successor of Alphadon. From Cox (1988). |
Marsupials were much more succesful on the southern continents. They arrived in South
America relatively late in the Cretaceous, but by early Paleocene time this group of
mammals was flourishing there. Marsupials make up more than 50% of the mammal species in
some South American Paleocene faunas, and adaptive types include insectivores, omnivores,
carnivores and small herbivores. Paleocene mammals are not yet known from Antarctica and
Australia. However, these continents were connected to each other and to South America at
that time, and Eocene marsupials occur there. So it is highly probable that marsupials
lived there during the Paleocene as well.
Many Paleocene marsupials of South America are closely related to todays opossums,
which are appropriately called "living fossils". Opossums and their relatives
belong to the order Didelphimorphia (named after the recent Virginia opossum Didelphis).
Pucadelphys, a primitive opossum from the early Paleocene of Bolivia, is known from
the best fossils for any early marsupial. Nearly complete skeletons have been found in
pairs, which are interpreted as male and female. They were probably trapped in their
burrows during a flood. Pucadelphys was an agile animal with some capabilities for
bounding and digging, but without strong adaptations for climbing in trees. Like most
recent opossums it was probably an omnivore that fed on insects, fruits and small
vertebrates.
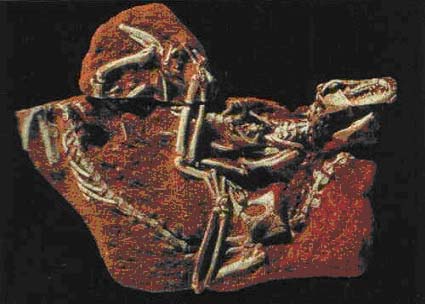 |
| Figure 6: Pair of skeletons of the opossum Pucadelphys andinus
from the early Paleocene of Bolivia, preserved in a life-like way just as resting in their
nest. Skeleton on top of the figure faces to the right, skeleton at the bottom (without
skull) to the left. From Rich & Rich (1993). |
Abundant fossil Didelphimorphia are known from middle Paleocene fissure fillings in
Brazil. Members of this order cover a large range of size and adaptation at that time. Minusculodelphis
was close in size to the smallest known mammals. It is based on fossil jaws that contain
teeth less than a millimeter in size! This tiny animal probably hunted for insects. Eobrasilia
was much larger and had bulbous teeth similar to Late Cretaceous Didelphodon. It
was previously considered as a relative of the carnivorous Sparassodonta discussed below,
but is now regarded as an independent branch of opossum relatives that evolved towards a
more carnivorous niche.
A second group of marsupials, the order Paucituberculata, is known in South America
from the Paleocene to the present. During the Paleocene the most important members were
the extinct polydolopids, rat to rabbit-sized animals that were somewhat similar to
rodents in ecology. The middle Paleocene genus Epidolops is known from a nearly
complete skull. All other polydolopids are only known from fossil jaws and teeth. Like
rodents, polydolopids had enlarged front teeth. Still more distinctive was that some of
their cheek teeth, especially the last premolars, formed large blades for cutting food.
Similar blade-like cheek teeth have evolved independently in several groups of mammals,
for instance the Mesozoic to early Tertiary multituberculates and todays rat-kangoroos.
Such teeth could be useful for processing tough parts of plants, but an animal with this
dentition could be an omnivore as well. Polydolopids declined in diversity during the
Eocene and became extinct in the Oligocene, probably due to competition with other groups
of Paucituberculata and with immigrating rodents and primates.
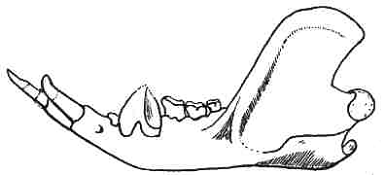 |
Figure 7: Lower jaw of the polydolopid Epidolops ameghinoi
from the middle Paleocene of Brazil. Note the large front teeth and the blade-like last
premolar. Later polydolopids like Polydolops developed strong serrations on this
tooth. From Paula Couto (1952). |
Finally, a group of marsupials started during the Paleocene to occupy the ecological
role of carnivorous mammals in South America. These animals are classified in the extinct
order Sparassodonta, which culminated in the dog- or bear-like borhyaenids and the
saber-toothed cat-like thylacosmilids of the later Cenozoic. Paleocene members of this
order demonstrate the transition from primitive opossum-like animals such as Pucadelphys
to ferocious predators. Mayulestes from the early Paleocene of Bolivia was a
small predator which occupied a ecological niche close to that of weasels or martens.
Adaptations like a prehensile tail show that this agile animal lived partially in trees.
On the ground Mayulestes was probably capable of relatively fast but short runs to
catch its prey.
The first larger sparassodonts are known from the middle Paleocene of Brazil. They
include the medium sized genus Patene (meaning 'fox' in a native South American
language) and a poorly known wolf-sized predator. These marsupial carnivores developed
basically the same tooth structure for cutting meat as the true (placental) carnivores of
the northern hemisphere. Unlike true carnivores, however, sparassodonts did not develop
particular adaptations for fast running. This may have been one reason for their decline
and finally their extinction after the arrival of true carnivores in South America in late
Cenozoic time.
Sources of figures:
Cox, B. (editor) 1988: Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals.
Macmillan London Limited.
Miao, D. (1993): Cranial morphology and multituberculate melationships. In: Szalay et.
al. (editors), Mammal Phylogeny, volume 1. Springer-Verlag New York, Inc.
Paula Couto, C. de 1952: Fossil mammals from the beginning of the Cenozoic in Brazil.
Marsupialia: Polydolopidae and Borhyaenidae. Amer. Mus. Novitates 1559, 1-27.
Rich, P. V. & Rich, T. H. 1993: Wildlife of Gondwana. Dinosaurs and other
verterbates from the ancient supercontinent.
Scillato-Yané, G. J. & Pascul, R. (1985): Un peculiar Xenarthra del paleoceno
medio de Patagonia (Argentina). Su importancia en la sistemática de los Paratheria.
Ameghiniana 21, 173-176
Sloan, R. E. (1981): Systematics of Paleocene multituberculates from the San Juan
Basin, New Mexico. In: Lucas et. al. (editors), Advances in San Juan Basin Paleontology,
University of New Mexico Press.
Stanley, S. M. 1989: Earth and Life Through Time. Freeman and Company.